Histamine Intolerance: The Complete Clinical Guide
Symptoms, Genetics, Receptors, Associated Conditions and Evidence-Based Strategies
By Alex Manos MSc (Nutrition) | IFM-Certified Functional Medicine Practitioner
Do you experience headaches after a glass of wine, skin flushing after eating leftovers, or persistent digestive symptoms that no-one can explain? You may be dealing with histamine intolerance — a clinically under-recognised condition rooted in the biochemistry of histamine metabolism, receptor biology, and gut health. This guide covers everything you need to know about histamine: what it is, how it is degraded, the genetics that make some people vulnerable, and the evidence-based strategies that can help.
1. What Is Histamine?
Histamine is a biogenic amine synthesised from the amino acid L-histidine by the enzyme histidine decarboxylase (HDC). It is stored primarily in mast cells and basophils and is released in response to immune stimulation, injury, allergens, and certain foods. Once released, it acts as a potent local signalling molecule, a neurotransmitter in the brain, and a key mediator of the immune system (Maintz & Novak, 2007).
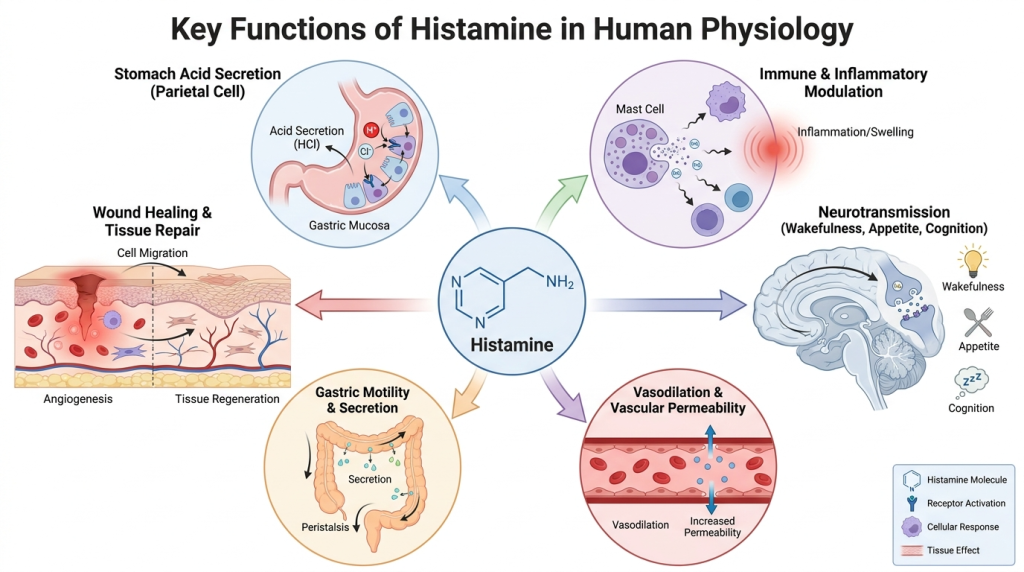
Histamine is found throughout the body, including in the gut, skin, lungs, cardiovascular system, and brain. It plays physiologically vital roles including:
- Regulating stomach acid secretion via parietal cells
- Modulating immune and inflammatory responses
- Acting as a neurotransmitter influencing wakefulness, appetite, and cognition
- Facilitating vasodilation and vascular permeability
- Contributing to gastric motility and intestinal secretion
- Playing a role in wound healing and tissue repair
The challenge arises not from histamine itself, but from the body’s ability to keep it in balance. When histamine accumulates beyond the body’s capacity to break it down, a spectrum of symptoms — collectively referred to as histamine intolerance — can emerge.
2. Where Does Histamine Come From?
Histamine in the body arises from two broad sources:
2.1 Endogenous Histamine
The body produces histamine via mast cells, basophils, enterochromaffin-like (ECL) cells in the gastric mucosa, platelets, and neurons in the hypothalamus. In the CNS, histaminergic neurons originate in the tuberomammillary nucleus of the hypothalamus and project widely throughout the brain (Haas & Panula, 2003).
2.2 Exogenous Histamine — Food Sources
Dietary histamine is the primary exogenous source. Histamine accumulates in foods that have undergone fermentation, ageing, or microbial breakdown. High-histamine foods include:
- Aged and fermented cheeses (parmesan, camembert, cheddar)
- Cured and processed meats (salami, pepperoni, bacon)
- Fermented foods (sauerkraut, kimchi, kombucha, vinegar)
- Alcohol — particularly red wine, champagne, and beer
- Smoked and tinned fish
- Tomatoes, spinach, and aubergine
- Leftovers (histamine increases with storage time)
Beyond direct histamine content, some foods act as histamine liberators – substances that trigger mast cell degranulation and histamine release even when they do not contain histamine themselves. These include strawberries, citrus fruits, pineapple, papaya, peanuts, and shellfish. Other foods, such as alcohol and black tea, inhibit the enzyme diamine oxidase (DAO), further impairing clearance (Maintz & Novak, 2007).
2.3 Gut Microbiome as a Source of Histamine
An often-overlooked source is the gut microbiome. Certain bacterial species possess histidine decarboxylase and can convert dietary histidine to histamine directly in the gastrointestinal tract. A 2021 genomic study published in BMC Genomics identified histamine-producing bacteria across multiple phyla, including Proteobacteria, Firmicutes, Fusobacteriota, and Verrucomicrobiota (Mou et al., 2021).
A 2022 study published in Nutrients examining patients with histamine intolerance found significantly higher abundance of histamine-secreting bacteria — including genera such as Staphylococcus, Proteus, Clostridium perfringens, and Enterococcus faecalis — compared to healthy controls, alongside a notably lower proportion of beneficial taxa such as Faecalibacterium prausnitzii and Ruminococcus (Comas-Basté et al., 2022).
This microbiome dimension means that histamine intolerance is not solely a question of dietary intake or enzyme deficiency — the composition of the gut microbiota itself can be a significant driver of histamine load.
3. How Is Histamine Metabolised?
The body relies on two primary enzymatic pathways to clear histamine:
3.1 Diamine Oxidase (DAO)
DAO (diamine oxidase) — encoded by the AOC1 gene — is the principal enzyme responsible for degrading extracellular, exogenous histamine in the gastrointestinal tract. It is expressed primarily by mature enterocytes in the intestinal mucosa, and it oxidatively deaminates histamine, converting it to imidazole acetaldehyde. DAO activity is highest in the small intestinal villi, making it the first line of defence against dietary histamine absorption (Maintz & Novak, 2007).
DAO requires copper and vitamin B6 (pyridoxal-5′-phosphate) as cofactors. It is also critically dependent on adequate vitamin C, which supports DAO enzyme function. Numerous factors reduce DAO activity, including gut inflammation, intestinal permeability (leaky gut), coeliac disease, inflammatory bowel disease, SIBO, alcohol, and specific medications including non-steroidal anti-inflammatory drugs (NSAIDs), certain antibiotics, and proton pump inhibitors (Hrubisko et al., 2021).
3.2 Histamine N-Methyltransferase (HNMT)
HNMT is the intracellular enzyme responsible for methylating and thereby inactivating histamine within cells, primarily in the central nervous system, liver, and kidneys. Unlike DAO, which works in the gut lumen, HNMT operates at the cellular level. It requires S-adenosyl methionine (SAMe) as a methyl donor — a cofactor directly dependent on efficient methylation pathways (Horton et al., 2005).
HNMT is the dominant histamine-degrading pathway in the brain. When HNMT activity is impaired, intracellular histamine accumulates in neuronal tissue, potentially contributing to neurological symptoms such as anxiety, brain fog, insomnia, and headaches. The product of HNMT metabolism — N-methylhistamine — is subsequently broken down by monoamine oxidase B (MAO-B).
4. The Genetics of Histamine Intolerance
Why do some people accumulate histamine while others tolerate the same dietary load without symptoms? A significant part of the answer lies in genetic variation. Single nucleotide polymorphisms (SNPs) in the genes encoding histamine-degrading enzymes and related methylation pathways can substantially reduce enzymatic capacity.
4.1 AOC1 Gene (DAO Enzyme)
The AOC1 gene encodes the DAO enzyme. Several functionally significant SNPs have been identified in this gene, including rs10156191, rs2052129, and rs1049742 (Maintz et al., 2011). These variants are associated with reduced serum DAO activity, which correlates with increased symptom severity. A 2015 study published in Headache found that specific AOC1 polymorphisms (rs10156191 and rs2052129) were significantly associated with migraine risk — a clinical finding consistent with the role of impaired histamine clearance in migraine pathophysiology (García-Martín et al., 2015).
The AOC1 gene is located on chromosome 7q35-36. DAO activity can vary considerably between individuals based on their genetic profile, and this variation helps explain why two individuals eating identical diets may have completely different symptom profiles.
4.2 HNMT Gene
The HNMT gene contains well-characterised polymorphisms that reduce intracellular histamine methylation. The most studied is the C314T (Thr105Ile) polymorphism (rs11558538), which results in an amino acid substitution that substantially reduces HNMT enzyme activity. The T allele at this position is linked to impaired intracellular histamine clearance (Preuss et al., 1998).
Another functional SNP is T939C (rs1050891) in the 3′ untranslated region, which influences HNMT mRNA stability and consequently enzyme expression levels. Given that HNMT is the primary CNS pathway for histamine clearance, HNMT polymorphisms are likely to be of particular significance in individuals with predominantly neurological or psychiatric histamine symptoms — including anxiety, depression, ADHD, and sleep disturbance (Heidari et al., 2015).
4.3 MTHFR Gene
The MTHFR (methylenetetrahydrofolate reductase) gene encodes the enzyme that converts 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate — a critical step in the methylation cycle that ultimately produces SAMe. Since HNMT requires SAMe as its methyl donor, impaired MTHFR function reduces the availability of SAMe and consequently reduces HNMT-mediated histamine clearance.
The most clinically relevant MTHFR variants are C677T (rs1801133) and A1298C (rs1801131). The C677T homozygous variant can reduce MTHFR enzyme activity by approximately 70%, while the heterozygous variant reduces activity by approximately 35% (Frosst et al., 1995). The A1298C variant is also associated with reduced BH4 (tetrahydrobiopterin) synthesis, which may contribute to increased mast cell degranulation and histamine release (Fryar-Williams, 2016).
Individuals with MTHFR variants may therefore struggle with histamine clearance not just because their HNMT enzyme is compromised, but because impaired methylation has downstream effects on neurotransmitter balance, detoxification, and immune regulation — all of which interact with the histamine system.
4.4 HDC Gene (Histidine Decarboxylase)
The HDC gene encodes histidine decarboxylase, the enzyme that produces histamine from L-histidine. Gain-of-function variants in HDC increase histamine synthesis and have been implicated in Tourette syndrome, ADHD, and other histaminergic disorders. While less commonly discussed in the context of dietary histamine intolerance, variations in HDC can contribute to excessive endogenous histamine production (Castellan Baldan et al., 2014).
Key clinical insight: Histamine intolerance is rarely due to a single gene. Most patients have a combination of genetic vulnerabilities — often a DAO SNP increasing gut histamine load, an HNMT or MTHFR SNP reducing clearance, and gut dysbiosis increasing histamine production. This multi-factorial model explains why one-dimensional approaches (such as diet alone) often fail to resolve symptoms completely.
5. Histamine Receptors: Why Symptoms Affect So Many Body Systems
One of the most important concepts for understanding the diverse symptom picture of histamine intolerance is receptor biology. Histamine does not act uniformly throughout the body — its effects depend entirely on which receptor it binds to, and each receptor subtype is distributed differently across tissues. There are four known histamine receptor subtypes: H1, H2, H3, and H4. All four are G protein-coupled receptors (GPCRs) with characteristic 7-transmembrane domain architecture (Seldeslachts et al., 2023).
5.1 H1 Receptor — The ‘Classic’ Allergy Receptor
The H1 receptor is primarily expressed in smooth muscle, endothelium, neurons, and immune cells. It mediates the most familiar histamine effects and is the target of classic antihistamine medications. H1 receptor activation produces:
- Bronchoconstriction (relevant in asthma and allergic rhinitis)
- Vasodilation and increased vascular permeability — causing flushing, urticaria, and oedema
- Pruritus (itching) — via stimulation of sensory nerve endings
- Modulation of wakefulness and circadian rhythm in the CNS
- Smooth muscle contraction — contributing to intestinal cramping and motility changes
H1 activation is also involved in the sensation of pruritus in combination with H4 receptors (Wilzopolski et al., 2021). This explains why H1 antihistamines alone do not always abolish itching in certain inflammatory skin conditions.
5.2 H2 Receptor — Gastric and Cardiac Effects
The H2 receptor is densely expressed in gastric parietal cells, the cardiovascular system, and immune cells. H2 activation is responsible for:
- Stimulating gastric acid secretion — the basis of H2-blocker therapy (e.g., famotidine) in GORD and peptic ulcer disease
- Positive inotropic and chronotropic effects on the heart (increased heart rate and force)
- Modulation of gastrointestinal motility and intestinal secretion
- Downregulation of certain immune responses (immunomodulation)
In histamine intolerance, H2-mediated gastric acid hypersecretion can mimic or exacerbate GORD, causing heartburn, reflux, and acid-related symptoms. H2 receptor activation in cardiac tissue explains why some individuals with high histamine loads experience palpitations, tachycardia, and blood pressure fluctuations (Del Valle, 2006).
5.3 H3 Receptor — Brain and Nervous System Regulation
The H3 receptor acts as a presynaptic autoreceptor and heteroreceptor, primarily in the central and peripheral nervous system. It functions as a negative feedback mechanism, inhibiting histamine synthesis and release when activated. H3 activation also modulates the release of other neurotransmitters including dopamine, serotonin, noradrenaline, and acetylcholine. Key effects include:
- Regulation of appetite and energy balance
- Modulation of sleep-wake cycles and arousal
- Involvement in cognition, memory, and attention
- Regulation of pain perception
- Association with Tourette syndrome pathophysiology (Pittenger, 2020)
The H3 receptor’s role in neurotransmitter regulation means that dysregulated histamine signalling can have broad neuropsychiatric consequences — including effects on mood, concentration, and sleep quality.
5.4 H4 Receptor — Immune and Inflammatory Signalling
The H4 receptor was the last to be characterised and is principally expressed in bone marrow, peripheral leukocytes, mast cells, eosinophils, and the small intestine. It is now recognised as a key mediator of inflammatory and immune processes. H4 activation is associated with:
- Recruitment and activation of eosinophils and mast cells
- Proinflammatory cytokine production
- Contribution to pruritus alongside H1 (Wilzopolski et al., 2021)
- Potential involvement in inflammatory bowel disease and atopic dermatitis
- Possible roles in tumour microenvironment regulation
No H4 receptor antagonist has yet received clinical approval as of 2025, but preclinical studies have shown promise in asthma, dermatitis, and inflammatory bowel models (NCBI StatPearls, 2025).
Clinical implication: The diversity of histamine receptor distribution explains why histamine intolerance can present so differently between individuals. A person with predominantly H1-driven symptoms may present with urticaria and nasal congestion. Another with H2 dominance may primarily suffer reflux and palpitations. A third with H3-predominant effects may experience brain fog, insomnia, and anxiety. Recognising which receptor subtypes appear most active clinically can help guide targeted interventions.
5.5 Receptor Distribution Summary
- H1: Smooth muscle, endothelium, neurons → Flushing, bronchospasm, itch, urticaria, vasodilation
- H2: Gastric parietal cells, heart, immune cells → Acid hypersecretion, palpitations, tachycardia, immunomodulation
- H3: CNS neurons (presynaptic) → Brain fog, insomnia, mood changes, appetite dysregulation
- H4: Immune cells, bone marrow, gut → Inflammation, immune activation, eosinophilia, itch
6. What Is Histamine Intolerance?
Histamine intolerance (HIT) is defined as a disequilibrium between histamine intake and/or endogenous production, and the body’s capacity to metabolise and degrade it. The result is an accumulation of histamine in the circulation and tissues that provokes a range of pseudo-allergic symptoms (Maintz & Novak, 2007).
It is important to distinguish histamine intolerance from:
- IgE-mediated food allergy — which involves an immune response and can cause anaphylaxis
- Mast Cell Activation Syndrome (MCAS) — which involves dysregulated mast cell degranulation and excess release of multiple mediators, not just histamine
- Mastocytosis — a clonal mast cell disorder involving mast cell proliferation
Histamine intolerance is primarily a metabolic phenomenon — a mismatch between load and clearance — rather than an immune or structural condition. However, MCAS and histamine intolerance can co-exist, and MCAS should be considered when symptoms are disproportionately severe, multi-system, or triggered by non-dietary stimuli such as stress, exercise, heat, and hormonal fluctuation.
6.1 Prevalence
Histamine intolerance is estimated to affect approximately 1–3% of the general population, with women considerably more affected than men. This sex difference is significant and relates directly to the hormonal modulation of histamine metabolism discussed in Section 8 (Maintz & Novak, 2007; Hrubisko et al., 2021).
6.2 The Symptom Bucket Concept
A useful clinical model is the ‘histamine bucket’. Each individual has a threshold for histamine tolerance. When the cumulative load — from dietary intake, gut bacterial production, endogenous release, and impaired degradation — exceeds that threshold, symptoms spill over. This explains why symptoms are often inconsistent: a patient may tolerate a glass of wine on a low-stress day but react badly to the same quantity after a stressful week, when gut health is poor, or during the high-oestrogen phase of the menstrual cycle.
7. The Symptom Spectrum of Histamine Intolerance
Because histamine receptors are distributed across virtually every organ system, the symptom picture of histamine intolerance is notoriously broad and can mimic many other conditions. This is one reason why HIT is so frequently underdiagnosed or misattributed.
7.1 Gastrointestinal Symptoms
- Bloating and postprandial fullness
- Abdominal cramping and pain
- Diarrhoea and/or constipation
- Nausea and vomiting
- Acid reflux and heartburn (H2-mediated)
- Loose stools after high-histamine meals

7.2 Neurological and Cognitive Symptoms
- Headaches and migraines (one of the most common presentations)
- Brain fog and impaired concentration
- Anxiety and irritability
- Insomnia and sleep disruption
- Dizziness and vertigo
- Fatigue unrelated to physical exertion
7.3 Dermatological Symptoms
- Urticaria (hives) — acute or chronic
- Flushing — particularly of the face, neck, and chest
- Pruritus (itching) without visible rash
- Eczema exacerbations
- Angioedema (swelling of subcutaneous tissue)
7.4 Cardiovascular Symptoms
- Heart palpitations and tachycardia (H2-mediated)
- Hypotension — particularly postprandial
- Flushing and visible vasodilation
7.5 Respiratory Symptoms
- Rhinorrhoea (runny nose) and nasal congestion
- Sneezing
- Bronchospasm in susceptible individuals
- Asthma exacerbation
7.6 Menstrual and Hormonal Symptoms
- Worsening of symptoms around ovulation and pre-menstrually
- Dysmenorrhoea (painful periods)
- Premenstrual syndrome and PMDD symptom overlap
- Endometriosis symptom exacerbation
8. Conditions Associated with Histamine Dysregulation
Histamine dysregulation does not exist in isolation. It is now recognised as a contributing mechanism in a growing number of chronic conditions. Understanding this context is essential for effective clinical management.
8.1 Migraine
The relationship between histamine and migraine is one of the most thoroughly documented. Histamine provokes vasodilation of cranial vasculature via H1 receptors, and low DAO activity has been consistently associated with increased migraine frequency. A randomised controlled trial published in Clinical Nutrition found that DAO supplementation significantly reduced migraine frequency in patients with confirmed DAO deficiency (Izquierdo-Casas et al., 2019). Specific AOC1 gene polymorphisms have been identified as risk factors for dietary migraine (García-Martín et al., 2015). Additionally, meningeal mast cells are in close proximity to cranial sensory receptors and release histamine and other inflammatory mediators during migraine attacks (Conti et al., 2019).
8.2 Irritable Bowel Syndrome (IBS)
There is substantial evidence linking histamine excess to IBS. Mast cell density is elevated in the intestinal mucosa of IBS patients, and histamine release from these cells activates intestinal sensory neurons and modulates motility and secretion. Analysis of a US database including over 53 million patients found that individuals with IBS were at least four times more likely to have a mast cell disorder compared to the general population (Kurin et al., 2023). Gut dysbiosis with histamine-producing bacteria further compounds the burden in IBS patients, and a low-histamine diet has been shown to reduce IBS symptom scores in clinical practice.
8.3 Mast Cell Activation Syndrome (MCAS)
MCAS is characterised by recurrent episodes of mast cell mediator release affecting two or more organ systems. While histamine intolerance results primarily from impaired degradation, MCAS involves dysregulated mast cell activation. The 2022 updated consensus criteria require documentation of symptomatic mast cell mediator release alongside elevated mediators such as serum tryptase (20% + 2 ng/mL above baseline) or elevated urinary N-methylhistamine, prostaglandin D2, or leukotriene E4. Importantly, MCAS and histamine intolerance frequently co-exist, and the distinction between them requires careful clinical and laboratory assessment.
8.4 Chronic Urticaria
Both acute and chronic spontaneous urticaria are strongly associated with histamine release. A randomised, double-blind, placebo-controlled trial published in the International Archives of Allergy and Immunology found that DAO supplementation significantly reduced symptom burden in chronic spontaneous urticaria patients compared to placebo (Yacoub et al., 2018). Mast cell degranulation — whether IgE-mediated or IgE-independent — drives histamine-dependent whealing and flare responses.
8.5 Atopic Dermatitis and Eczema
Atopic dermatitis involves complex H1 and H4 receptor-mediated itch signalling. H4 receptor activation recruits eosinophils and mast cells to the skin, sustaining inflammatory cycles. Elevated histamine levels and dysregulated mast cell function are consistently found in atopic dermatitis patients, and low-histamine dietary approaches show supportive benefit in some subgroups.
8.6 Fibromyalgia
Emerging clinical evidence links histamine intolerance and DAO deficiency to fibromyalgia. A double-blind, placebo-controlled trial found that DAO supplementation significantly reduced pain in fibromyalgia patients with confirmed DAO deficiency, suggesting histamine-mediated central sensitisation as a potential mechanistic pathway (Okutan et al., 2023).
8.7 Anxiety, Depression, and Psychiatric Symptoms
Histamine is an important neurotransmitter in the central nervous system, with H3 receptors regulating the release of dopamine, serotonin, and noradrenaline. Dysregulated histaminergic neurotransmission has been implicated in depression, anxiety, and schizophrenia (Qian et al., 2022). HNMT polymorphisms that impair intracellular histamine methylation may contribute to neurological symptom profiles. Notably, mutations in HNMT have been associated with autosomal recessive intellectual disability, underscoring the neurodevelopmental importance of HNMT function (Heidari et al., 2015).
8.8 SIBO and Small Intestinal Dysbiosis
Small intestinal bacterial overgrowth (SIBO) can significantly impair histamine metabolism. Bacterial overgrowth in the small intestine both increases local histamine production (via histamine-producing species) and drives mucosal inflammation that reduces DAO expression in enterocytes. Conversely, histamine-driven gut dysmotility may predispose to SIBO — creating a bidirectional relationship. Approximately 31% of patients with MCAS have been found to have concurrent SIBO (Kresser Institute, 2026).
8.9 Hormonal Conditions — Endometriosis, PMDD, and Oestrogen Excess
The relationship between oestrogen and histamine is bidirectional and clinically significant. Oestrogen stimulates mast cell degranulation and histamine release, while simultaneously downregulating DAO activity. Histamine, in turn, stimulates ovarian granulosa cells to produce more oestrogen via H1 receptors — creating an amplifying cycle (Verhoeven, 1993). Progesterone has the opposing effect: it stabilises mast cells and upregulates DAO, which explains why many women experience worsening histamine symptoms at ovulation and pre-menstrually (when progesterone is falling), and improvement during the luteal phase when progesterone is at its highest.
9. Diagnosing Histamine Intolerance
Diagnosing histamine intolerance remains challenging due to the absence of a single validated biomarker and the breadth of overlapping symptom presentations. A systematic diagnostic approach is essential.
9.1 Serum DAO Activity
Measurement of serum DAO activity is currently the most widely used diagnostic marker. A retrospective cohort study of 192 patients with histamine intolerance symptoms published in Nutrients (2022) found that symptom severity correlated significantly with the degree of DAO deficiency, and that patients with intermediate DAO levels (3–10 U/mL) showed the greatest clinical improvement on a low-histamine diet and/or DAO supplementation (Cucca et al., 2022). Mušič et al. (2013) demonstrated significantly lower DAO activity in histamine intolerance patients compared to healthy controls, with symptoms resolving after histamine-free diet in patients with highly reduced DAO activity (<40 HDU/mL).
It is important to note that serum DAO activity does not perfectly reflect intestinal mucosal DAO activity, and a normal DAO level does not rule out histamine intolerance — particularly in individuals with HNMT or MTHFR polymorphisms causing impaired intracellular clearance.
9.2 Urinary N-Methylhistamine
24-hour urinary N-methylhistamine (NMH) is a metabolite of HNMT-mediated histamine breakdown and serves as a marker of overall histamine turnover. Elevated NMH suggests increased histamine load or impaired clearance and is used as a supportive diagnostic marker, particularly in the assessment of MCAS alongside prostaglandin D2 and leukotriene E4.
9.3 Elimination and Reintroduction Diet
A structured low-histamine elimination diet for 4–8 weeks followed by systematic histamine food reintroduction remains the most clinically practical diagnostic tool. Symptom resolution on elimination and recurrence on reintroduction is the diagnostic gold standard in the absence of validated biomarkers. Compliance and dietary education are essential, as patients often inadvertently include high-histamine foods.
9.4 Genetic Testing
Genetic testing for AOC1 (DAO), HNMT, and MTHFR polymorphisms via platforms such as 23andMe or clinical SNP panels can identify genetic vulnerability but should be interpreted alongside clinical context — the presence of a SNP does not confirm active disease, and conversely, symptom response to a low-histamine diet can occur in the absence of known genetic variants.
10. Evidence-Based Management Strategies
Management of histamine intolerance is most effective when individualised and multi-pronged, addressing the key drivers: dietary load, enzymatic capacity, gut health, stress, and hormonal balance.

10.1 Dietary Management
Low-Histamine Diet
A low-histamine diet eliminates or substantially reduces foods high in histamine, histamine liberators, and DAO inhibitors. Foods to avoid include aged cheeses, cured and fermented meats, fermented vegetables, tinned and smoked fish, alcohol, vinegar-containing foods, tomatoes, spinach, aubergine, strawberries, citrus fruits, and leftovers. Fresh, minimally processed foods cooked and eaten promptly are the foundation of the diet.
A clinical study published in Nutrients demonstrated that patients with confirmed HIT experienced significant symptom improvement on a low-histamine diet and that adherence correlated with a measurable increase in serum DAO activity (Schnedl et al., 2019). A 2024 review confirmed the central role of dietary management in HIT, noting that the combination of dietary restriction and DAO supplementation provides superior outcomes to either alone (Comas-Basté et al., 2024).
Practical Dietary Principles
- Cook from scratch using fresh ingredients
- Eat leftovers on the same day or freeze immediately after cooking
- Prioritise low-histamine protein sources: fresh poultry, beef, and freshly caught fish
- Favour tolerated vegetables: courgette, broccoli, cauliflower, leafy greens (except spinach)
- Avoid prolonged marinating, which increases histamine content
- Be cautious with restaurant meals — fermentation and ageing processes are difficult to control
Important: The low-histamine diet is an investigative and therapeutic tool, not a permanent prescription. The goal is to reduce overall histamine load, allow DAO activity to recover, and progressively reintroduce foods to establish individual tolerance thresholds.
10.2 Gut Health Optimisation
Given the central role of gut inflammation in reducing DAO activity, and of gut dysbiosis in increasing histamine production, restoring gut health is foundational to long-term histamine management.
Addressing Dysbiosis
A pilot study published in Nutrients (2022) demonstrated that a low-histamine diet combined with DAO supplementation reduced histamine-secreting bacteria — specifically Proteus and Raoultella — and increased Roseburia species over 9 months, demonstrating that dietary treatment actively modulates the microbiome towards a lower-histamine state (Sánchez-Pérez et al., 2022).
Probiotic Selection
Probiotic choice matters significantly in histamine intolerance, as several commonly used strains — including some Lactobacillus casei, Lactobacillus bulgaricus, and Lactobacillus helveticus strains — are histamine producers. Clinically preferred strains include:
- Lactobacillus rhamnosus (LGG) — well studied for gut barrier support, does not produce histamine
- Bifidobacterium infantis — gut barrier support, anti-inflammatory properties
- Bifidobacterium longum — histamine-degrading capacity demonstrated in vitro
- Lactiplantibacillus plantarum — identified as non-histamine-producing with strong mucosal adhesion (Russo et al., 2025)
Leaky Gut and Mucosal Repair
Intestinal hyperpermeability allows histamine to cross the mucosal barrier and enter systemic circulation more readily. Supporting mucosal integrity through adequate zinc, vitamin D, glutamine, and butyrate-producing fibre intake creates a structural basis for improved histamine management.
10.3 Supplement Support
DAO Enzyme Supplementation
Oral DAO supplementation provides exogenous enzyme activity in the gastrointestinal tract, supporting histamine degradation of dietary histamine when taken before meals. An open-label interventional pilot study of 28 patients with confirmed HIT published in Nutrients found significant improvement in GI symptoms — including bloating, abdominal pain, and diarrhoea — during the DAO supplementation phase, with symptoms returning when supplementation was withdrawn (Schnedl et al., 2019). Clinical recommendations suggest DAO supplementation is taken 15–20 minutes before high-histamine meals.
Vitamin C
Vitamin C (ascorbic acid) has a well-documented antihistamine effect, reducing serum histamine levels and supporting DAO enzyme function. A clinical study found that intravenous vitamin C significantly decreased serum histamine concentrations in patients with allergic and non-allergic diseases (Hagel et al., 2013). Vitamin C should be sourced from non-citrus, low-histamine sources where possible — for example, acerola cherry extract, rosehip, or buffered ascorbate preparations. Typical supportive dosing ranges from 500–2,000 mg daily.
Quercetin
Quercetin is a plant polyphenol flavonoid that acts as a natural mast cell stabiliser, inhibiting histamine release from activated mast cells. It also inhibits histidine decarboxylase (thereby reducing histamine synthesis) and provides anti-inflammatory effects. Clinical evidence demonstrates quercetin’s capacity to reduce allergic responses via mast cell stabilisation (Mlcek et al., 2016). Quercetin’s bioavailability is enhanced by bromelain co-administration. Typical clinical dosing ranges from 500–1,000 mg daily with food.
Vitamin B6 (P5P)
Pyridoxal-5′-phosphate (P5P), the active form of vitamin B6, is an essential cofactor for the DAO enzyme. Suboptimal B6 status directly impairs DAO activity. P5P supplementation at doses of 25–50 mg daily is a rational support strategy for individuals with confirmed DAO deficiency, particularly those with poor dietary B6 intake or absorption.
Vitamin B12 and Methylfolate
For individuals with MTHFR variants, supporting the methylation cycle with methylated B vitamins — specifically methylcobalamin (B12) and 5-methyltetrahydrofolate (5-MTHF) — supports SAMe production, which is required as the cofactor for HNMT-mediated histamine clearance. This is particularly relevant for patients with neurological histamine symptoms including anxiety, brain fog, and insomnia.
Copper
DAO is a copper-dependent enzyme. Clinical copper deficiency is uncommon, but marginal status could contribute to reduced DAO activity. Dietary copper sources include liver, shellfish, nuts, and seeds. Where copper supplementation is considered, the zinc:copper ratio should be monitored, as high-dose zinc supplementation can deplete copper.
Luteolin
Luteolin is a flavonoid with potent mast cell-stabilising properties. Research demonstrated that luteolin inhibits mast cell secretion and reduces neuroinflammation, suggesting particular relevance for neurological histamine symptoms including brain fog and anxiety (Theoharides et al., 2015). Luteolin is found in celery, green peppers, parsley, and chamomile, and is available in supplemental form.
10.4 Stress Management
Psychological stress is a significant and clinically underappreciated driver of histamine release. Stress stimulates the release of corticotropin-releasing hormone (CRH) from the hypothalamus, which directly activates mast cells expressing CRHR-1 receptors to degranulate and release histamine and other inflammatory mediators. A comprehensive review published in the Annals of Allergy, Asthma & Immunology concluded that psychological stress worsens many histamine-driven conditions — including asthma, atopic dermatitis, and mastocytosis — through this CRH-mast cell pathway (Theoharides et al., 2020).
Chronic stress additionally dysregulates the HPA axis and reduces cortisol’s anti-inflammatory buffering capacity, tipping the balance towards mast cell activation. This creates a physiologically important link between chronic psychological stress and histamine symptom burden — one that dietary interventions alone cannot address.
Practical Stress Management Strategies
- Daily mindfulness or meditation practice — even 10 minutes has measurable HPA axis regulatory effects
- Diaphragmatic breathing and vagal nerve activation to reduce mast cell activation via the autonomic nervous system
- Adequate and consistent sleep — circadian disruption increases mast cell reactivity
- Regular moderate exercise — while high-intensity exercise can transiently elevate histamine, consistent moderate activity reduces chronic inflammatory tone
- Psychological support or therapy — particularly for individuals with identifiable chronic stressors
- Social connection and nature exposure — evidence-based downregulators of the stress response
10.5 Hormonal Considerations
Given the oestrogen-histamine feedback loop, hormonal balance is an important clinical variable — particularly in women. Strategies to support healthy oestrogen metabolism include:
- Optimising the oestrobolome — the gut microbial genes involved in oestrogen conjugation and recirculation — through fibre, diverse plant foods, and microbial diversity
- Supporting liver phase I and II oestrogen detoxification through adequate B vitamins, cruciferous vegetables (as tolerated), and calcium-D-glucarate
- Considering natural progesterone support in consultation with a clinician where progesterone deficiency is confirmed — progesterone stabilises mast cells and upregulates DAO
- Minimising xenoestrogen exposure from plastics, personal care products, and conventional meat to reduce oestrogen burden
11. The Gut-Histamine Axis: A Functional Medicine Perspective
From a functional medicine perspective, the gut is the central interface of histamine biology. It is where the majority of exogenous histamine is introduced, where DAO is produced and deployed, where histamine-producing bacteria reside, and where gut inflammation can compromise all of these processes simultaneously.
A functional approach to histamine intolerance therefore begins with a comprehensive gut assessment: microbiome composition and histamine-producing bacterial load, intestinal permeability markers, stool inflammation markers, SIBO testing where clinically indicated, and DAO serum activity. Treatment follows the 5R framework applied specifically to histamine drivers:
Remove: High-histamine foods, histamine-producing bacterial overgrowth, and triggers of mast cell activation
Replace: DAO enzyme supplementation, digestive support, and cofactors (vitamin B6, vitamin C, copper)
Reinoculate: Low-histamine probiotic strains and prebiotic fibre to restore beneficial microbial populations
Repair: Mucosal integrity through zinc, glutamine, vitamin D, and butyrate support
Rebalance: Stress management, hormonal balance, sleep, and circadian rhythm — the upstream regulators of mast cell reactivity
12. Summary: Key Mechanisms and Interventions
Histamine load too high? → Low-histamine diet, address gut dysbiosis, treat SIBO if present
DAO deficiency (genetic or acquired)? → DAO supplementation, vitamin B6, vitamin C, copper repletion
HNMT/MTHFR impairment? → Methylated B vitamins (methylcobalamin, 5-MTHF), reduce intracellular histamine load
Gut dysbiosis with histamine-producing bacteria? → Low-histamine probiotics, dietary treatment, antimicrobial support as indicated
Mast cell hyperreactivity (MCAS co-existing)? → Quercetin, luteolin, vitamin C, stress management, hormonal balance, and specialist assessment
Oestrogen-histamine cycle? → Oestrogen metabolism support, progesterone balance review, oestrobolome optimisation
Chronic stress as a driver? → HPA axis support, mindfulness, vagal toning, sleep hygiene
13. Conclusion
Histamine is far more than a molecule associated with hay fever and hives. It is a ubiquitous signalling molecule whose actions — mediated through four distinct receptor subtypes distributed across virtually every tissue in the body — encompass neurotransmission, immune regulation, gastric secretion, cardiovascular function, and reproductive biology.
Histamine intolerance emerges at the intersection of genetic vulnerability, enzymatic capacity, gut microbiome composition, dietary load, stress physiology, and hormonal status. Its symptom picture is broad because its receptor biology is broad. This complexity is a clinical challenge, but also an opportunity: addressing the root causes of histamine accumulation — rather than simply blocking receptors with antihistamines — offers the prospect of genuine, durable resolution.
An evidence-based functional medicine approach, integrating low-histamine dietary principles, gut microbiome restoration, targeted supplementation, stress management, and hormonal balance, provides a comprehensive framework for managing even complex, multi-system histamine presentations.
If you are struggling with unexplained chronic symptoms that you suspect may be histamine-related, a comprehensive assessment including serum DAO activity, gut microbiome analysis, and genetic testing can help identify the root cause and guide a personalised treatment plan. Visit alexmanos.co.uk to explore gut microbiome testing, SIBO breath testing, and functional medicine consultations.
References
Castellan Baldan, L., Williams, K.A., Gallezot, J.D., et al. (2014). Histidine decarboxylase deficiency causes Tourette syndrome: parallel findings in humans and mice. Neuron, 81(1), 77–90. https://doi.org/10.1016/j.neuron.2013.10.052
Comas-Basté, O., Sánchez-Pérez, S., Veciana-Nogués, M.T., Latorre-Moratalla, M., & Vidal-Carou, M.C. (2020). Histamine intolerance: the current state of the art. Biomolecules, 10(8), 1181. https://doi.org/10.3390/biom10081181
Comas-Basté, O., Sánchez-Pérez, S., Latorre-Moratalla, M.L., & Vidal-Carou, M.C. (2022). Intestinal dysbiosis in patients with histamine intolerance. Nutrients, 14(9), 1774. https://doi.org/10.3390/nu14091774
Conti, P., D’Ovidio, C., Conti, C., et al. (2019). Progression in migraine: role of mast cells and pro-inflammatory and anti-inflammatory cytokines. European Journal of Pharmacology, 844, 87–94. https://doi.org/10.1016/j.ejphar.2018.12.004
Cucca, V., Ramirez, G.A., Pignatti, P., et al. (2022). Basal serum diamine oxidase levels as a biomarker of histamine intolerance: a retrospective cohort study. Nutrients, 14(7), 1513. https://doi.org/10.3390/nu14071513
Del Valle, J. (2006). Zollinger-Ellison syndrome. In: Sleisenger and Fordtran’s Gastrointestinal and Liver Disease, 8th edn. Saunders Elsevier.
Fiorani, M., Del Vecchio, L.E., Dargenio, P., et al. (2023). Histamine-producing bacteria and their role in gastrointestinal disorders. Expert Review of Gastroenterology & Hepatology, 17(7), 709–718. https://doi.org/10.1080/17474124.2023.2230865
Frosst, P., Blom, H.J., Milos, R., et al. (1995). A candidate genetic risk factor for vascular disease: a common mutation in methylenetetrahydrofolate reductase. Nature Genetics, 10(1), 111–113. https://doi.org/10.1038/ng0595-111
Fryar-Williams, S. (2016). Fundamental role of methylenetetrahydrofolate reductase 677 C→T genotype and flavin compounds in biochemical phenotypes for schizophrenia and schizoaffective psychosis. Frontiers in Psychiatry, 7, 172. https://doi.org/10.3389/fpsyt.2016.00172
García-Martín, E., Martínez, C., Serrador, M., et al. (2015). Diamine oxidase rs10156191 and rs2052129 variants are associated with the risk for migraine. Headache, 55(2), 276–286. https://doi.org/10.1111/head.12493
Haas, H.L., & Panula, P. (2003). The role of histamine and the tuberomamillary nucleus in the nervous system. Nature Reviews Neuroscience, 4(2), 121–130. https://doi.org/10.1038/nrn1034
Hagel, A.F., Layritz, C.M., Hagel, W.H., et al. (2013). Intravenous infusion of ascorbic acid decreases serum histamine concentrations in patients with allergic and non-allergic diseases. Naunyn-Schmiedeberg’s Archives of Pharmacology, 386(9), 789–793. https://doi.org/10.1007/s00210-013-0880-1
Heidari, A., Tongsook, C., Najafipour, R., et al. (2015). Mutations in the histamine N-methyltransferase gene, HNMT, are associated with nonsyndromic autosomal recessive intellectual disability. Human Molecular Genetics, 24(20), 5697–5710. https://doi.org/10.1093/hmg/ddv286
Horton, J.R., Sawada, K., Nishibori, M., et al. (2005). Two polymorphic forms of human histamine methyltransferase: structural, thermal, and kinetic comparisons. Structure, 13(11), 1645–1655. https://doi.org/10.1016/j.str.2005.07.019
Hrubisko, M., Danis, R., Huorka, M., & Wawruch, M. (2021). Histamine intolerance — the more we know the less we know. A review. Nutrients, 13(7), 2228. https://doi.org/10.3390/nu13072228
Izquierdo-Casas, J., Comas-Basté, O., Latorre-Moratalla, M.L., et al. (2019). Diamine oxidase (DAO) supplement reduces headache in episodic migraine patients with DAO deficiency: a randomised double-blind trial. Clinical Nutrition, 38(1), 152–158. https://doi.org/10.1016/j.clnu.2018.01.013
Maintz, L., & Novak, N. (2007). Histamine and histamine intolerance. American Journal of Clinical Nutrition, 85(5), 1185–1196. https://doi.org/10.1093/ajcn/85.5.1185
Maintz, L., Yu, C.F., Rodríguez, E., et al. (2011). Association of single nucleotide polymorphisms in the diamine oxidase gene with diamine oxidase serum activities. Allergy, 66(7), 893–902. https://doi.org/10.1111/j.1398-9995.2011.02558.x
Mlcek, J., Jurikova, T., Skrovankova, S., & Sochor, J. (2016). Quercetin and its anti-allergic immune response. Molecules, 21(5), 623. https://doi.org/10.3390/molecules21050623
Mou, Z., Yang, Y., Hall, A.B., & Jiang, X. (2021). The taxonomic distribution of histamine-secreting bacteria in the human gut microbiome. BMC Genomics, 22, 695. https://doi.org/10.1186/s12864-021-08004-3
Mušič, E., Korošec, P., Šilar, M., Adamič, K., Košnik, M., & Rijavec, M. (2013). Serum diamine oxidase activity as a diagnostic test for histamine intolerance. Wiener Klinische Wochenschrift, 125(9–10), 239–243. https://doi.org/10.1007/s00508-013-0354-y
Okutan, G., Tobajas, J., González-Colominas, E., et al. (2023). DAO supplementation in fibromyalgia: a double-blind placebo-controlled clinical trial. Journal of Clinical Medicine, 12(20), 6449. https://doi.org/10.3390/jcm12206449
Pittenger, C. (2020). The histidine decarboxylase model of tic pathophysiology: a new focus on the histamine H3 receptor. British Journal of Pharmacology, 177(3), 570–579. https://doi.org/10.1111/bph.14606
Preuss, C.V., Wood, T.C., Szumlanski, C.L., et al. (1998). Human histamine N-methyltransferase pharmacogenetics: common genetic polymorphisms that alter activity. Molecular Pharmacology, 53(4), 708–717. https://doi.org/10.1124/mol.53.4.708
Qian, H., Shu, C., Xiao, L., & Wang, G. (2022). Histamine and histamine receptors: roles in major depressive disorder. Frontiers in Psychiatry, 13, 825591. https://doi.org/10.3389/fpsyt.2022.825591
Russo, P., Florio, C., De Salvo, E., et al. (2025). Potential role of probiotic strain Lactiplantibacillus plantarum in control of histamine metabolism. Microorganisms, 13(6), 1299. https://doi.org/10.3390/microorganisms13061299
Sánchez-Pérez, S., Comas-Basté, O., Veciana-Nogués, M.T., Latorre-Moratalla, M.L., & Vidal-Carou, M.C. (2022). The dietary treatment of histamine intolerance reduces the abundance of some histamine-secreting bacteria of the gut microbiota in histamine intolerant women. Nutrients, 14(22), 4821. https://doi.org/10.3390/nu14224821
Schnedl, W.J., Lackner, S., Enko, D., et al. (2019). Diamine oxidase supplementation improves symptoms in patients with histamine intolerance. Food Science and Biotechnology, 28(6), 1779–1784. https://doi.org/10.1007/s10068-019-00627-3
Seldeslachts, A., Peigneur, S., & Tytgat, J. (2023). Histamine receptors: ex vivo functional studies enabling the discovery of hits and pathways. Membranes, 13(12), 897. https://doi.org/10.3390/membranes13120897
Theoharides, T.C., Kavalioti, M., & Taliou, A. (2015). Luteolin, a phytochemical with diverse biological activities and its structure-activity relationships. Frontiers in Pharmacology, 6, 295. https://doi.org/10.3389/fphar.2015.00295
Theoharides, T.C., Tsilioni, I., & Hua, J. (2020). The impact of psychological stress on mast cells. Annals of Allergy, Asthma & Immunology, 125(4), 388–392. https://doi.org/10.1016/j.anai.2020.07.007
Verhoeven, T.S. (1993). The effect of histamine on progesterone and estradiol secretion of human granulosa cells in serum-free culture. Human Reproduction, 8(11), 1965–1968. https://doi.org/10.1093/oxfordjournals.humrep.a137966
Wilzopolski, J., Kietzmann, M., Mishra, S.K., Stark, H., Bäumer, W., & Rossbach, K. (2021). Histamine H1, H3 and H4 receptors are involved in pruritus. Biomolecules, 11(8), 1166. https://doi.org/10.3390/biom11081166
Yacoub, M.R., Ramirez, G.A., Berti, A., et al. (2018). Diamine oxidase supplementation in chronic spontaneous urticaria: a randomised, double-blind placebo-controlled study. International Archives of Allergy and Immunology, 176(3–4), 268–271. https://doi.org/10.1159/000488142